










2012
May
20
The Bacterial Iron War Front
Well, here it is. My friends out there, knowing I'm a chemist, had to suppose that fairly soon there would be something here about chemistry. I am a physical/computational chemist and this post is more about biochemistry, but biochemistry needs computations, too. This particular topic relates to some literature research I am doing at the university right now.
You know that your body needs iron. It is part of what makes up hemoglobin, which transports oxygen to your cells. Without it, you die. Fewer people know that it is also a component of myoglobin, the primary oxygen-carrying species in muscle tissue. There is a large number of iron-sulfur proteins that perform crucial catalytic functions also necessary to keep you alive. In short, iron has quite a number of essential biological functions.
And it's not just humans. Bacteriophages use iron in the tips of their tail to attach successfully to a host cell, such as E. coli. E.coli, in turn, as do other bacteria, for its own growth and reproduction. This need for iron all over the place turns into a nanoscopic battle that our macroscopic eyes tend to miss.
The Subterfuge: Stealing Supplies
The first plan would be for bacteria to simply absorb iron from its environment, and in the case of a virulent E. coli infection, that environment might be you. Stealing your own iron to use it against you might not sound ethical, but bacteria aren't much into ethics, and it makes good military sense. Unfortunately for the bacteria, hosts have planned for that very contingency.
The First Line of Defense: Iron Starvation
The ferric (Iron(III) for you chemists) ion is relatively insoluble in the first place. Furthermore, most of that iron is kept locked up in the aforementioned hemoglobin, myoglobin, iron-sulfur proteins, and other bound states, safely inside cells where bacteria can't get at it. Still, there is a little bit floating around. Enter the transferrins. These are proteins whose job it is to grab hold of those free ferric ions in serum so that bacteria can't make use of it. Human transferrin has a binding constant of about 1023 , and only about 30% of it is actually bound, leaving a lot more to keep things swept up. If you crunch the numbers, this works out to only about four ions per septillion (US usage) left free in the serum. With odds like that, it sounds as if we have those nasty germs starved, doesn't it? Wrong!
The First Wave: Siderophores
Not willing to give up without a fight, bacteria produce siderophores in low-iron environments such as you. These are small
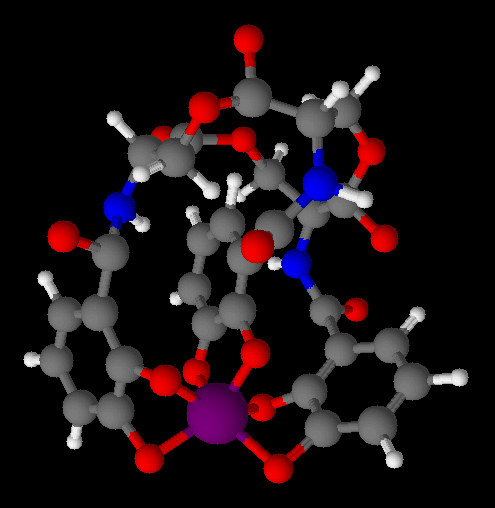
molecules who sole purpose it is to scavenge iron and take it back to the cell. One of the most common of these is enterobactin, and we'll assume we're talking about that one from now on. There might be only four ions per septillion free, but the enterobactin will find them, and when it does, the chemical equilibrium will shift to the left, causing the transferrin to release a few more ions. Fortunately for you, and unfortunately for the bacteria, this is a diffusion limited process. In other words, the enterobactin has to wander around until it actually bumps into one of those few iron ions, and then find its way back to the cell.
But of course, it's not that simple. The iron-enterobactin (FeEnt) binding constant is about 1049 , which is a whole lot bigger than iron-transferrin. It is only willing to leave just 4 ions out of 1050 free to roam around. Yikes! This means that, depending on the kinetic energy barrier, enterobactin and other siderophores might be able to simply steal iron from ferritin ... and indeed, it turns out that it can.
The Second Line of Defense: Interception
Of course, all the iron-enterobactin in the world won't do the germ any good wandering around in the plasma; it has to get back into the cell somehow. Cell membranes are composed, essentially, of fat, which repels ions of all kinds. So to get them inside, the cell expresses proteins that form ion channels designed to conduct an ion or ionophore through the nonpolar barrier. The first-line ion channel to get FeEnt through the bacteria's outer membrane is designated FepA.
Fortunately, it seems our bodies know all about that. There are mechanism in place to intercept that iron-loaded enterobactin before it can get to the FepA. One of the primary players in this defense is believed to be neutrophil gelatinase-associated lipocalin (NGAL). It has an affinity for FeEnt comparable to that of FepA, so if it gets to the enterobactin before the enterobactin finds its way back to the bacterium, it just never gets there.
The Second Wave: Flank Attack
Not surprisingly, the old germ has this base covered, too. If it can keep the whole battle for iron close to home territory, then NGAL won't have much of a chance to intercept it. Indeed, Simonson and others discovered that in time of iron stress, at least some bacteria express a protein that adheres to the cell surface and tightly binds transferrin. This allows enterobactin to pick it to the bone and make it into the cell before NGAL can run into it. This particular study did not identify siderophores as the active mechanism of iron extraction in this case, but that strategy will be assisted by this process as well as any other possible strategies.
The Third Line of Defense: Disguise
The host counters this strategy by expressing an overabundance of transferrin. With only 30% of transferrin carrying iron, that leaves a lot of iron-free transferrin (also called apotransferrin) on the prowl. The bacterium is unable to distinguish between iron-transferrin and apotransferrin, and so binds the latter just as enthusiastically as it does the former, only it does it no good. The energy and material used to produce the binding protein has been wasted, and the binding site is now clogged with something that to the bacterium's point of view is worthless.
The Backup Plan: Secret Weapon
While the host has been focused on the enterobactin route, blocking and counterblocking, the bacteria have been developing their secret weapon. It has been found out relatively recently that many bacteria can obtain iron through another route entirely: that of iron citrate. This method gets back to being diffusion limited and dependent on the amount of free iron as we started out with at the beginning, but it represents an entirely different route that bypasses the enterobactin scenario completely. Bacteria, like investors, know not to put all their eggs in one basket.
This is as far as I'll take the adventure, because here we are getting away from my research area. But I hope I have impressed upon you that this whole battle over iron involves dozens of specialized proteins representing tens of thousands of DNA codons. God or nature, depending on your world view, has assembled a system of startling complexity over the singular matter of obtaining and controlling ferric ions, a degree of complexity that does represent two seasoned generals pondering over their maps or two chess masters pondering over their board.
And this is just one detail among the thousands that permit a living organism to exist. As a matter of fact, it is just one detail among the hundreds that define the relationship between host and infectious organism. Just wait until you see how viruses incite sedition and rebellion, provoking your own cells to work against you and assist the enemy.
We know many aspects of biological systems to this level of detail. We have the human genome decoded. But there are still thousands of pieces missing, and it even when they are all found, it will be a long time before we will be able to assemble them into a coherent whole. Interested in biochemistry? Never fear; there is still plenty of work to do.
Aisen, Leibman, and Zweier, Stoichiometric and Site Characteristics for Binding of Iron to Human Transferrin
Raymond, Dertz, and Kim, Enterobactin: An archetype for microbial iron transport
Loomis and Raymond, Solution equilibria of enterobactin and metal-enterobactin complexes
Carrano and Raymond, Ferric Ion Sequestering Agents. 2. Kinetics and Mechanism of Iron Removal from Transferrin by Enterobactin and Synthetic Tricatecholsl
Simonson, Brener, and DeVoe, Expression of a High-Affinity Mechanism for Acquisition of Transferrin Iron by Neisseria meningitidis
Lensbouer, Patel, Sirianni, and Doyle, Functional Characterization and Metal Ion Specificity of the Metal-Citrate Complex Transporter from Streptomyces coelicolor




Comments
There are no comments for this post.
You must be logged in to post a comment.